Fossilbevis og evolusjon
(oversatt fra 'The Evolution Revolution' Kap. 1-slutten -av Dr. Lee M. Spetner, Jerusalem, Israel, Juni 2014.)
Om jeg ville holde noen kaniner innestengt i et bur, og etter noe tid fant flere kaniner i buret, så kunne jeg være sikker på at de tidligste kaninene ga opphav til de som kom senere. Om fossilfunn forekommer i gelogogiske lag, oppå hverandre, og de ser mer eller mindre like ut, så kan jeg med rimelig sikkerhet slutte at de i øvre lag er avkom etter de i nedre lag, men så sikker som med kaninene kunne en ikke være. Om de i øvre lag er noe ulike dem i nedre lag, så kunne jeg ennå helle mot at de nedre ga opphav til de øvre. Men likevel er det grunn til seriøse reservasjoner mot formodningen. I et slikt tilfelle har paleontologene N. Eldridge og S. Gould (1972) foreslått at de øvre ofte ikke var født på det stedet, men kan ha flyttet inn fra et annet territorium. Så formodningen om at fossiler i øvre lag, nedstammer fra de i lavere lag, er ikke nødvendigvis den eneste fortolkningen av fossile data.
Likesom logikken i forbindelse med fylogenetiske stamtre var baklengs, så er også logikken for å slutte fra fossiler til felles avstamning bakvendt. Om felles avstamning er sant, så kan en forvente å finne fossiler som viser stadier i evolusjon. Men en kan ikke nødvendigvis slutte andre veien. Det meste en kan si, er at fossilene ville være konsistente med evolusjons fortellingen. G. Himmelfarb sier det slik i forbindelse med fossiler og evolusjon: "Hver paleontologisk oppdagelse, som syne å ha evolusjonær betydning, blir på en eller annen måte tatt som bekreftelse på teorien om naturlig seleksjon, selv om de ikke har den fjerneste tilknytning til den teorien (Himmelfarb, 1962, 446).
Bilde 1. En Ikke-Tilfeldig Evolusjons-Hypotese
Imidlertid kan fossilene gjøres rede for ved EITEH (En Ikke Tilfeldig Evolusjons Hypotese), som vi snart skal se eks. på. Endringer fra ett geologisk lag til ett annet, kan godt skyldes den type endringer (lenke). De eks. innebar ingen gevinst av genetisk informasjon, for enten var de resultat av innebygde program eller de nye genene var allerede i populasjonen. Dermed blir ikke fossilene noe bevis for tillagt informasjon. Ikke i noe tilfeller er i så fall informasjon lagt til biosfæren, så evolusjonære endringer kan ikke låne støtte til felles avstamning. En teori om felles avstamning må gjøre rede for oppbygning av ny informasjon {- i og med at genom i høyere dyreordener vanligvis er mer omfattende enn i laverestående-oversetters tilføyelse}.
Når darwinister skal begrunne felles avstamning ut fra fossil-registeret, så benytter de ofte teologisk heller enn vitenskapelig begrunnelse. Argumentet kan gå på at Skaperen ikke ville legg fossiler i jorda for å forvirre oss. På det grunnlag slutter de at det ikke var noen skapelse og at hypotesen om felles avstamning må være sann. Forfatterne er ikke teologer, likevel har de dumdristighet til å forvirre med teologiske argumenter. Her vil vi bare komme med vitenskapelige bevis for mangelen på bevis for felles avstamning. Når det gjelder teologisk begrunnelse, henvises til dette eller dette. Dette nevnes bare for å få en kontrast til teologiske argument fra darwinister.
Bilde 2. Ida-funnet førte til noen overdrevne reaksjoner
 Darwinistiske forfattere refererer til fossiler, som de kaller overgangsformer som bevis for felles avstamning (6). Men fossilene har ingen teori å knytte seg til. Lignende argumenter ut fra fossil-arkivet kunne framsettes ut fra skapelse. Den slags bevis er ikke vitenskapelige. Darwinister henfører til fossiler som de hevder er mellomformer (se IDA). De påstår at disse fossiler er levninger av dyr som var i en overgangsfase fra en hovedform til en annen. Dette eks. fikk ny vind i seilene, på grunn av nylig funn av Tiktaalik fossilet (Daeschler et al.,2006). Fossilet er blitt hyllet som en missing link, som løser evolusjonsmysteriet -The Guardian 6.apil 1986). Det er det siste funnet mellom fisk og amfibier (Rogers 2011, Coyne 2009) og påstås å være videre bevis for evolusjon (felles avstamning).
Darwinistiske forfattere refererer til fossiler, som de kaller overgangsformer som bevis for felles avstamning (6). Men fossilene har ingen teori å knytte seg til. Lignende argumenter ut fra fossil-arkivet kunne framsettes ut fra skapelse. Den slags bevis er ikke vitenskapelige. Darwinister henfører til fossiler som de hevder er mellomformer (se IDA). De påstår at disse fossiler er levninger av dyr som var i en overgangsfase fra en hovedform til en annen. Dette eks. fikk ny vind i seilene, på grunn av nylig funn av Tiktaalik fossilet (Daeschler et al.,2006). Fossilet er blitt hyllet som en missing link, som løser evolusjonsmysteriet -The Guardian 6.apil 1986). Det er det siste funnet mellom fisk og amfibier (Rogers 2011, Coyne 2009) og påstås å være videre bevis for evolusjon (felles avstamning).
Fossiler fra fem utdødde dyr har blitt plassert på den evolusjonære linja fra fisk til tetrapods (firbente dyr). Fra den eldste til den yngste er de: Eusthenopteron, Panderichtlys, Tiktaalik, Acanthostega og Ichtyostega. I tråd med det neo-darwinistiske paradigmet er de plasser i en orden som skal illustrere en progresjon i evolusjonære overganger, fra fisk til firbente dyr. Framdriften er selvsagt hevdet ut fra formodningen om felles avstamning, så det kan knapt kalles bevis for felles avstamning. Andre fortolkninger er mulig her, som andre steder. Faktisk kan de også våre mer fornuftige. Fortolkningen om felles avstamning er i et hvert tilfelle sannsynligvis usann, siden tilfeldige mutasjoner og naturlig seleksjon ikke kan gjøre rede for den.
Noe kan også sees i lys av den innebygde kapasiteten til dyr til å tilpasse seg ved å svare på input fra omgivelsene. I følge datering av disse fossilene, er disse artene satt til å være omtrent i samme tidsperiode. Alle er datert til Sen-Devon periode. Estimerte datoer spenner over en tidsperiode på ca. 25 mill. år og varierer fra 385 mill. år til for ca. 360 mill. år siden. Men fossilspor fra en tetrapod (firfotet dyr), som skulle være senere enn alle de fem nevnte, er funnet i Polen og datert til ca. 397 mill. år siden (Niedzwiedzki et al, 2010), som er tidligere enn noen av dem. Dateringen indikerer at disse artene skulle betraktes som samtidige, særlig når en tar i betraktning at få fossiler som foreløbig er funnet, har bidratt til datering. De fem nevnte artene ovenfor, kan godt være utspring fra de samme artene, som tilpasser seg til nye betingelser i omgivelsene ut fra EITEH (En Ikke Tilfeldig Evolusjons Hypotese).
R. Carroll (1995) en framtredende paleontolog ved McGill Universitet, har uttalt i forbindelse med formodede overganger fra vann til land-dyr: "trekk ved skjelettet som en gang ble ment å være unike for landdyr, dukket opp i en følge av dyr knyttet til vann, vann og landlige omgivelser, land og vann-miljø og halvt-landlige virveldyr, mens trekk som vanligvis ble forbundet med fisk, ble bevart i amfibier, Fordelingen av primitive og oppnådde egenskaper varierer fra livsforløps-linje til linje , og viser at mange trekk ble oppnådd eller tapt konvergent (forfatters uthevelse).
Carrol sier altså her at noen trekk som ble knyttet kun til landdyr, er funnet i flere av fiskeartene, og noen trekk som bare ble knyttet til fisk, er funnet igjen i landdyr. For å gjøre rede for disse fossilene med blandede egenskaper, så trekker darwinister fram konvergent evolusjon. Forfatteren her (Lee Spetner), foreslår heller at trekk som sluttes fra disse fossilene heller er resultat av en innebygd tilpasset reaksjon på endrede omgivelser, etter EITEH (En Ikke Tilfeldig Evolusjons Hypotese). De nevnte dyr i en formodet overgangssekvens, har kanskje ikke stammet fra hverandre i det hele, men kan ha kommet fra samme opprinnelige populasjon, og simultant ha blitt tilpasset varierende omgivelses-nisjer. Både darwinisters og forfatterens forslag er spekulasjoner, fordi vi omtaler begivenheter som fant sted i fortiden og som vi ikke har eksperimentell tilgang til.
Det er altså to mulige forklaringer, forfatterens og darwinisters: a) En mulig forklaring innbefatter konvergent evolusjon, basert på tilfeldige mutasjoner i dyrets DNA, en forestilling som ikke støttes av observasjoner, og ikke har teretisk oppbakking (darwinistisk forklaring) og b) Innebygd evne i dyr til å tilpasse seg nye omgivelser, som støtter seg til fenomen som observeres i dag (forfatterens fortolkning).

Bilde 3. Påvirkes sterkt av miljømessige endringer: Pupfish i Death Valley reservat
Felles avstamnings-forestillingen har intet monopol i å fortolke fossil-dataene. Det finnes bedre, og fossil-registeret støtter ikke entydig felles avstamning. R. Dawkins (2010) liker å sammenligne fortolkning av sekundære bevis med måten en detektiv fortolker bevis i en kriminalsak. Om sekundære bevis skal være relevante i en kriminalsak, må de være knyttet til en teori om hvordan forbrytelsen ble begått. Men om den teorien ikke er realiserbar, så kan ikke bevisene godkjennes. Fossiler kaster ikke lys over hvordan evolusjon kan tilføye/danne ny informasjon. Det må en teori om felles avstamning kunne adressere. Men det kan verken neo-darwinisme eller noen annen teori gjøre pr. i dag. En må derfor konkludere at fossilene ikke støtter felles avstamning.
Geografisk fordeling
Geografisk fordeling av planter og dyr er et annet darwinistisk argument for felles avstamning, basert på teologi og ikke på vitenskap. I to foregående avsnitt (her) og her har forfatteren (Lee Spetner) vist at dyr og planter har en innebygd kapasitet til langvarig tilpasning til nye omgivelser. Ett stimuli fra en ny omgivelse kan forårsake at de endrer sitt genom, for å tilpasse seg til den omgivelsen. Ingen tilfeldige mutasjoner er påkrevd i den forbindelse. Forfatteren har gitt eks på arvelige endringer i planter og dyr som er fremkalt av omgivelsene. Levende organismer er skjenket en evne til å fungere effektivt i en variasjon av omgivelser. Ulike versjoner av samme dyr eller plante kan bli funnet i ulike steder på jorda, fordi hver har gjennomgått en arvelig endring stimulert av omgivelsene. Darwinister prøver å benytte disse biogeografiske data til å argumentere for evolusjon ved å påstå at en skaper ikke ville gjøre det på den måten. Jerry Coyne skrev om planter: "Hvorfor skulle en skaper sette fundamentalt ulike planter, men ser så like ut i ulike områder av verden som synes økologisk identiske? Ville det ikke være mer fornuftig å sette samme plantearter i områder med samme type jordsmonn og klima (Coyne, 2009, s.91)"
Om dyr skrev han: "Om dyr var spesielt skapt, hvorfor skulle skaperen produsere fundamentalt ulike dyr på ulike kontinenter, som likevel ser og handler så mye likt? (Coyne, 2009, s.88)" Forfatteren (Lee Spetner) håper han leser denne boka. Hans formål er ikke bare å imøtegå argumentet, han ønsker å rasere argumentet fra biogeografi. R. Dawkins skrev: "Hvorfor skulle en allmektig skaper beslutte å sette sine omhyggelig dannede arter på øyer og kontinenter, i nøyaktig passende mønstre til uimotståelig å foreslå at de har utviklet seg og finfordelt seg fra deres utgangspunkt i evolusjonen? 'Mønsteret' foreslår ikke noe i den retning. Det er kun en darwinistisk automatikk-forslag.
Det Coyne og Dawkins sier her er at de 'vet' at tilfeldige mutasjoner og naturlig seleksjon kan produser ulike planter og at dyr med lignende karaktertrekk på ulike steder. Kreasjonist-alternativet, for dem, er at skaperen former atskilte planter og dyr på hver eneste lokasjon, og de betrakter de som absurd. Men Skaperen viser seg flinkere enn både Coyle og Dawkins. Han synes virkelig å ha omhyggelig dannet informasjon i sine arter, til å gi dem evne til å svare på omgivelsenes stimuli, for å endre sitt genom til nye omgivelser. Så lot ham dem vandre dit de ville, med en evne til å tilpasse seg. Det er formastelig av Coyne, Dawkins og deres like å trekke fram amatør-messige teologiske argumenter for å støtte hva de hevder er en vitenskapelig hypotese. Hele premisset for deres argument om evolusjon, basert på geografisk fordeling av planter og dyr er feilaktig og deres argument er ugyldig.
En kjede av tilpassede mutasjoner, som ikke støtter felles avstamning
 Om den darwinistiske historien var sann, så måtte det ha eksistert et stort antall av tilfeldige mutasjoner som hadde økt informasjon i gen-sammensetning, som tidligere ikke fantes i biosfæren. Disse mutasjonene måtte ha blitt bygd en på en, for å oppnå sekvensen av økende tilpasning. Om slike lange sekvenser av mutasjoner virkelig inntreffer, så skulle vi være i stand til å observere noen av dem i virksomhet nå. I det minste burde vi kunne finne fragmenter av slike inntreffende sekvenser, både i laboratoriet og i naturen. Vi burde også være i stand til vise hvordan slike fragmenter kunne utvides for å indikere resultatene til felles avstamning. Om en fant slike fragmenter, ville det gi litt troverdighet til felles avstamning. Om vi hadde et fragment av en sekvens på bare hundre av de mange millioner av tilpassede mutasjoner på mange ulike steder, så kunne vi ekstrapolere til de mange millioner som er nødvendig for felles avstamning. Men vi har ikke funnet en sekvens på 100, vi har ikke engang funnet en sekvens på 10, eller engang tre, som har økt informasjon til genomet. Vitenskapslitteraturen gir ikke eks. på én tilfeldig mutasjon som øker arvelig informasjon i genomet. {Vi kommenterer noen påstått slike nedenfor -oversetters merknad.} Dette er tunge bevis mot felles avstamning.
Om den darwinistiske historien var sann, så måtte det ha eksistert et stort antall av tilfeldige mutasjoner som hadde økt informasjon i gen-sammensetning, som tidligere ikke fantes i biosfæren. Disse mutasjonene måtte ha blitt bygd en på en, for å oppnå sekvensen av økende tilpasning. Om slike lange sekvenser av mutasjoner virkelig inntreffer, så skulle vi være i stand til å observere noen av dem i virksomhet nå. I det minste burde vi kunne finne fragmenter av slike inntreffende sekvenser, både i laboratoriet og i naturen. Vi burde også være i stand til vise hvordan slike fragmenter kunne utvides for å indikere resultatene til felles avstamning. Om en fant slike fragmenter, ville det gi litt troverdighet til felles avstamning. Om vi hadde et fragment av en sekvens på bare hundre av de mange millioner av tilpassede mutasjoner på mange ulike steder, så kunne vi ekstrapolere til de mange millioner som er nødvendig for felles avstamning. Men vi har ikke funnet en sekvens på 100, vi har ikke engang funnet en sekvens på 10, eller engang tre, som har økt informasjon til genomet. Vitenskapslitteraturen gir ikke eks. på én tilfeldig mutasjon som øker arvelig informasjon i genomet. {Vi kommenterer noen påstått slike nedenfor -oversetters merknad.} Dette er tunge bevis mot felles avstamning.
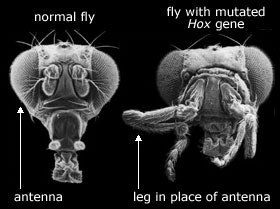
Bilde 4. Påstått 'suksessfull mutasjon': ben i stedet for antenne
Jerry Coyne (2009) skrev om et eksperiment som viste en sekvens av tre mutasjoner, som syntes å være hva forfatteren påsto ikke var funnet. Disse tre mutasjonene gir suksessivt en ny evne til en bakterie-populasjon. Coyne beskrev et eksperiment der et gen ble slettet fra E-coli bakteriet. Det genet kodet enzymet som startet nedbryting av laktose., og uten de kunne ikke bakterien ernære seg av laktose. Så ble bakterien med det manglende genet plassert i et miljø der laktose var eneste næring, slik at bakterien ikke kunne vokse. Men etter en tid dukket en mutasjon opp, som tillot dem å vokse sakte. Litt senere dukket en ny mutasjon opp, som gjorde veksten litt raskere. Enda senere dukket en tredje mutasjon opp, som tillot enda raskere vekst.
Tilsynelatende virker dette eksperimentet som det viser reell evolusjon. En kunne tro at disse tre mutasjonene, kunne være fragment av en lang sekvens. Coyne tilskrev eksperimentet til Barry Halls gruppe ved Universitetet i Rochester. Men Coyne hadde ikke spesifisert referansen. Hans referanseliste hadde bare en referanse til Hall alene fra 1982. I det eksperimentet ble et latent gen aktivert, som tok over jobben til det slettede genet. Hall beskriver ikke noe eksperiment der et gen ble slettet fra E-coli. Dette eksperimentet ble utført av andre, og var merkelig likt det Coyne refererte til. Halls artikkel fra 1982 ble utført på en jord-bakterie -diskutert i 'Not by Chance'. En populasjon av jordbakterier som normalt ernærte seg fra sukkertypen ribitol, ble plassert i en omgivelse som bare inneholdt sukkertypen xylitol, som bakterien ikke kunne benytte i utgangspunktet. Bakterien manglet enzym som kan bryte ned xylitol, , og de mangler enzym som kan transportere xylitol inn til cellen. Etter en stund inntraff imidlertid en mutasjon som tillot dem å vokse sakte på xylitol. Litt senere inntraff en en annen mutasjon som tillot dem å vokse litt raskere, og endelig en tredje mutasjon som tillot enda raskere vekst.
 Den første av de 3 mutasjonene var en punktmutasjon som deaktiverte repressoren til RDH-genet (7). Dette koder for RDH enzymet, som bryter ned ribitol. Å deaktivere repressoren, forårsaker RDH-enzym syntesen å gå for full fart, noe som gjør at store mengder av enzymet dannes. Selv om RDH-enzymet er spesifikt rettet mot ribito, så har det en liten virkning på xylitol(8). Mens denne aktiviteten vanligvis er for liten til at bakteriene kan leve på xylitol, så tillater det store antallet av enzym-molekylet en liten vekst i xylitol. Men fordi cellen ikke har noe transport-enzym for xylitol, så kan bare noen få molekyler komme inn i cellen ved diffusjon. Den tredje av mutasjonene, også en punktmutasjon, deaktiverte repressoren til transport-genet som koder for transport-enzymet til et annet sukker (D-arabitol). Det viser seg at dette viser seg å fungere bra på xylitol, så lenge D-arabitol finnes. På grunn av at det ble dannet mange transport-enzymer, ble mange xylitol molekyler tatt opp av cellen, noe som tillot raskere vekst.
Den første av de 3 mutasjonene var en punktmutasjon som deaktiverte repressoren til RDH-genet (7). Dette koder for RDH enzymet, som bryter ned ribitol. Å deaktivere repressoren, forårsaker RDH-enzym syntesen å gå for full fart, noe som gjør at store mengder av enzymet dannes. Selv om RDH-enzymet er spesifikt rettet mot ribito, så har det en liten virkning på xylitol(8). Mens denne aktiviteten vanligvis er for liten til at bakteriene kan leve på xylitol, så tillater det store antallet av enzym-molekylet en liten vekst i xylitol. Men fordi cellen ikke har noe transport-enzym for xylitol, så kan bare noen få molekyler komme inn i cellen ved diffusjon. Den tredje av mutasjonene, også en punktmutasjon, deaktiverte repressoren til transport-genet som koder for transport-enzymet til et annet sukker (D-arabitol). Det viser seg at dette viser seg å fungere bra på xylitol, så lenge D-arabitol finnes. På grunn av at det ble dannet mange transport-enzymer, ble mange xylitol molekyler tatt opp av cellen, noe som tillot raskere vekst.
Den andre mutasjonen var ikke i en repressor, men var en punktmutasjon i selve RDH-genet. Mutasjonen økte aktiviteten til RDH-enzymet i forhold til xylitol, og senket i forhold til ribitol. Alle disse kan få det til å se ut som informasjon vinnes, men nøyere informasjon viser at informasjon faktisk går tapt. Ved å teste dette modifiserte enzymet på et annet relatert sukker, viste at hva mutasjonen faktisk gjorde var å senke spesifisiteten til enzymet (flere detaljer i 'Not by Chance').
 Reduksjon i spesifisitet er tap av informasjon, akkurat som en adresse på en konvolutt 'Idrettssvingen, Sørum' inneholder mindre informasjon enn om det sto: 'Idrettssvingen 3, 1990 Sørum, Norge' fordi den første er mindre detaljert enn den andre. Videre må et enzym være spesifikt m.h.t. hvilken reaksjon det katalyserer. Spesifisiteten til et enzym er i hvert fall ikke mindre viktig enn nivået på aktiviteten. Et enzym som vil godta ethvert molekyl som sitt tilhørende substrat, kan virke skadelig. For det første sløser det ressurser ved overdreven bruk av energi og næring. Dessuten kan unødige produkter som det lager, blande seg med hva cellen holder på med. Som Bone, Silen og Agard (1989) skrev: "En av de fundamentale funksjonene til et enzym, er å forsyne spesifisitet ved å begrense området av substrater, som de er produktive i å katalysere." Det å miste spesifisiteten til enzymer, er ikke en måte å oppnå felles avstamning på. En repressor er et viktig element i cellekontroll. det sikrer at genet som kontrolleres er AV, når dets produkt ikke trengs. Det å deaktivere en repressor innebærer at genet er PÅ hele tiden. Selv om det kunne ha en positiv bieffekt i eksemplet ovenfor, så ville ikke cellen fungere bra under normale omstendigheter, med ikke-muterte celler. Flere mutasjoner av dette slaget, ville bare innebære mer deaktivering og mer degradering, som ikke er en formel for felles avstamning.
Reduksjon i spesifisitet er tap av informasjon, akkurat som en adresse på en konvolutt 'Idrettssvingen, Sørum' inneholder mindre informasjon enn om det sto: 'Idrettssvingen 3, 1990 Sørum, Norge' fordi den første er mindre detaljert enn den andre. Videre må et enzym være spesifikt m.h.t. hvilken reaksjon det katalyserer. Spesifisiteten til et enzym er i hvert fall ikke mindre viktig enn nivået på aktiviteten. Et enzym som vil godta ethvert molekyl som sitt tilhørende substrat, kan virke skadelig. For det første sløser det ressurser ved overdreven bruk av energi og næring. Dessuten kan unødige produkter som det lager, blande seg med hva cellen holder på med. Som Bone, Silen og Agard (1989) skrev: "En av de fundamentale funksjonene til et enzym, er å forsyne spesifisitet ved å begrense området av substrater, som de er produktive i å katalysere." Det å miste spesifisiteten til enzymer, er ikke en måte å oppnå felles avstamning på. En repressor er et viktig element i cellekontroll. det sikrer at genet som kontrolleres er AV, når dets produkt ikke trengs. Det å deaktivere en repressor innebærer at genet er PÅ hele tiden. Selv om det kunne ha en positiv bieffekt i eksemplet ovenfor, så ville ikke cellen fungere bra under normale omstendigheter, med ikke-muterte celler. Flere mutasjoner av dette slaget, ville bare innebære mer deaktivering og mer degradering, som ikke er en formel for felles avstamning.
Bilde 6. Spesifikk passform en forutsetning for proteiner/substrater
Ut fra å tilbakevise eneste påviste tilfeller av mutasjoner som kunne ha økt informasjonsinnhold i cellen, så kan det hevdes at ikke en eneste tilfeldig mutasjon som øker informasjonsinnholdet i cellen, er funnet.
Et annet forfeilet darwinist-argument
Darwin formodet at levende vesener konkurrerte sterkt mot hverandre, når deres antall økte høyere enn ressursene de ernærer seg på. Denne konkurransen fører i følge ham til naturlig seleksjon av de best tilpassede. Darwin overtok denne forestillingen fra Thomas Malthus (1798), som var bekymret for at populasjonen av mennesker til sist ville passere mattilgangen. Men om Malthus-spekulasjonen var sann om dyr, så burde vi se dyr som levde under miserable forhold, alltid på grensen til hungersnød. {Dette stemmer vanligvis bare under spesielle forhold, som tørkeperioder-oversetters kommentar.} Under slike harde forhold ville bare de tøffeste individene overleve og formere seg, i følge Darwin. Men her tok han feil.
 R. Dawkins prøver å omgjøre dette til et argument for evolusjon. Han benytter igjen et teologisk argument, om at hvis en Skaper hadde planlagt økosystemet, så ville det vært med tanke på velferd til helheten av aktuelle populasjoner. Under det darwinistiske paradigmet, der 'hvert individ er for seg selv', så ville byttedyr tendere i å overbeskatte sitt bytte. Han skriver: "skulle vi ikke forvente at ville rovdyr som ulver eller løver, var 'kloke rovdyr også'? Svaret er nei, nei.. Hans argument er at dette ville en skaper ha gjort, men han hevder at evolusjonen virker annerledes. Nå virker det som Dawkins for en gangs skyld benytter riktig teologi, men da er biologien han benytter feilaktig. Rovdyr overbeskatter generelt ikke populasjonen av byttedyr. Generelt har ulve- populasjoner en likevekt i forhold til og hjort/elg- populasjoner, som tenderer omkring likevekt (Messier og Crete, 1985, Messier 1994, Eberhardt og Peterson 1999, Eberhardt 2000, Messier og Joly 2000). Ulvene overbeskatter ikke hjort/elge- bestand, men begrenser i stedet sin egen populasjon for å sikre at hjort/elg fortsetter å fungere som mat for fremtidige ulvegenerasjoner (Wynne Edwards 1986, Bergerud 1983). De lever så å si bare på utbyttet av sine ressurser, og bevarer mesteparten for ettertiden. Til vanlig lever dyrepopulasjoner greit, noe som motsier at de oppfører seg som Dawkins hevder.
R. Dawkins prøver å omgjøre dette til et argument for evolusjon. Han benytter igjen et teologisk argument, om at hvis en Skaper hadde planlagt økosystemet, så ville det vært med tanke på velferd til helheten av aktuelle populasjoner. Under det darwinistiske paradigmet, der 'hvert individ er for seg selv', så ville byttedyr tendere i å overbeskatte sitt bytte. Han skriver: "skulle vi ikke forvente at ville rovdyr som ulver eller løver, var 'kloke rovdyr også'? Svaret er nei, nei.. Hans argument er at dette ville en skaper ha gjort, men han hevder at evolusjonen virker annerledes. Nå virker det som Dawkins for en gangs skyld benytter riktig teologi, men da er biologien han benytter feilaktig. Rovdyr overbeskatter generelt ikke populasjonen av byttedyr. Generelt har ulve- populasjoner en likevekt i forhold til og hjort/elg- populasjoner, som tenderer omkring likevekt (Messier og Crete, 1985, Messier 1994, Eberhardt og Peterson 1999, Eberhardt 2000, Messier og Joly 2000). Ulvene overbeskatter ikke hjort/elge- bestand, men begrenser i stedet sin egen populasjon for å sikre at hjort/elg fortsetter å fungere som mat for fremtidige ulvegenerasjoner (Wynne Edwards 1986, Bergerud 1983). De lever så å si bare på utbyttet av sine ressurser, og bevarer mesteparten for ettertiden. Til vanlig lever dyrepopulasjoner greit, noe som motsier at de oppfører seg som Dawkins hevder.
Bilde 7. Ulv og elge-populasjoner -se komm. i lenke
Wynne Edvards foreslo i 1986 at dyre populasjoner vanligvis holdes i sjakk, ikke av ytre krefter som sykdommer eller massedød, men av indre kontrollmekanismer bygd inn i dyrene. Selv om de fleste finner det overraskende, har det vært rapportert av biologer som studerer ville dyrebestander i flere ulike populasjoner. Her gikk R. Dawkins i ei felle: hans argument mot evolusjon, ble et argument mot den.
Stoffutvalg og bilder ved Asbjørn E. Lund